Высшие корковые функции человека и их нарушения при локальных поражениях мозга
Александр Романович Лурия
Этапы пройденного пути
Научная автобиография

|
Высшие корковые функции человека и их нарушения при локальных поражениях мозгаАлександр Романович ЛурияЭтапы пройденного пути
|
|
|
|
|
|||
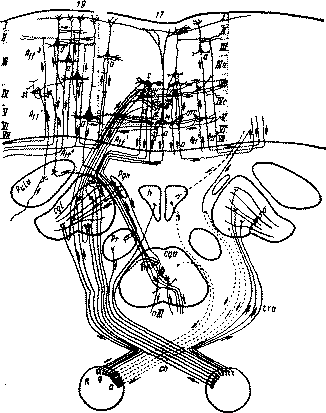
Высшие корковые функции человека и их нарушения при локальных поражениях мозгаб) Зрительный анализатор и строение зрительной корыОсновным препятствием для правильного понимания мозговых механизмов зрительного восприятия и для физиологической оценки явлений оптической агнозии была та «рецепторная» теория ощущений и восприятий, которая почти безраздельно господствовала в психологии и неврологии XIX и начала XX века и о которой мы частично уже говорили выше. Согласно этой теории, как это уже указывалось выше, ощущение является пассивным процессом, возникающим в результате раздражений органов чувств приходящими извне агентами. Возбуждения из сетчатки направляются к рецепторным центрам коры головного мозга, где они приобретают характер ощущений; лишь затем эти ощущения объединяются в восприятия, которые в свою очередь, перерабатываясь, превращаются в более 'сложные единицы познавательной деятельности. Первые этапы этого сложного пути «рецепторная» теория ощущений склонна была рассматривать как элементарные и пассивные физиологические процессы, последние этапы — как сложные и активные психические виды деятельности. Из этой теории неизбежно вытекал тот разрыв между элементарными и высшими формами познавательной деятельности, о котором речь шла выше. Иначе подходит ко многим из этих явлений рефлекторная концепция восприятия, основы которой были заложены еще И. М. Сеченовым и которая была экспериментально обоснована И. П. Павловым. В ряде психофизиологических исследований последнего времени (Гранит, 1956, Ε Η Соколов, 1958 и др.) эта теория была перенесена и «а исследование восприятия человека. Согласно рефлекторной теории ощущения и восприятия рассматриваются как активные процессы, отличающиеся известной избирательностью ,и включающие эфферентные (моторные) компоненты Физиологические механизмы этого избирательного анализа и синтеза раздражителей были подробно изучены И. П. Павловым и его сотрудниками. Этим была заложена основа физиологии анализаторов, осуществляющих избирательное выделение сигнальных (существенных для организма) и отвлечение от побочных (несущественных) признаков. Это и составило ядро рефлекторной теории восприятия. С точки зрения рефлекторной теории многие механизмы восприятия стали представляться существенно иначе. И. M. Сеченов указывал, что каждый акт зрительного восприятия включает в свой состав наряду с центростремительными (афферентными) еще и центробежные (эфферентные) механизмы Глаз, воспринимая предметы окружающего мира, активно «ощупывает» их, и эти «ощупывающие» движения наряду с проприоцептивными сигналами от глазодвигательных мышц входят как элементы в состав зрительного восприятия Открытие И M Сеченова оказалось исходным для большой серии работ, богато развернувшихся особенно в последние годы, после того, как новая техника позволила дать характеристику тех электрофизиологических явлений, которые протекают в отдельных участках зрительного анализатора, начиная с сетчатки глаза и кончая зрительной корой Оказалось, что процессы, происходящие на отдельных этапах зрительного пути, имеют не односторонний (центростремительный), а двусторонний (петально-фугальный или афферентно-эфферентный) характер и что наряду с разрядами, регистрируемыми в зрительном нерве и вышележащих центральных участках зрительного анализатора, при раздражении центральных отделов зрительного аппарата можно констатировать также изменения потенциалов сетчатки. Эти факты, отражающие явления, хорошо известные в психологии (например, повышение остроты зрения в зависимости от активного внимания или установки субъекта и τ д), были физиологически изучены Гранитом (1956), описавшим изменения потенциалов ретины под влиянием центральных воздействий, а у нас Ε Η. Соколовым (1958, 1959), проследившим роль ориентировочных реакций в протекании процессов восприятия и те сложные формы избирательных сензорных процессов, которые могут осуществляться лишь при ведущем участии коры. Эти исследования, указывающие на избирательный характер процессов восприятия, осуществляемых при участии системы двусторонних афферентно-эфферентных связей, и составляют фактическое обоснование рефлекторной теории ощущений. Какой же аппарат осуществляет сложные аналитико-синтетические процессы, лежащие в Основе зрительно-тактильных форм восприятия? Что известно нам о его строении и функциональных особенностях? Как мы должны представлять себе те нарушения воспринимающей деятельности, которые возникают при его поражении? Мы остановимся сначала в самых кратких чертах на том, что известно о строении зрительного анализатора, и особенно его корковых отделов. Строение зрительных отделов коры головного мозга, расположенных в пределах затылочной области, повторяет многое из того, что мы уже имели случай говорить, разбирая строение корковых отделов слухового анализатора. Мы напомним относящиеся сюда данные в самых общих чертах. Известно, что сетчатка глаза, в отличие от других периферических рецепторов, представляет собой сложнейшее нервное образование, рассматриваемое некоторыми авторами как кусочек коры головного мозга, вынесенный наружу. В ряде внутренних слоев сетчатки располагаются нейронные комплексы, одни из которых принимают возбуждения, возникающие в светочувствительных приборах наружного слоя, и передают их в изолированном или объединенном виде через волокна зрительного нерва на центральные нервные образования Другие нейронные группы принимают эфферентные импульсы, идущие от центральных образований, и служат аппаратами, которые осуществляют центральную регуляцию возбудимости отдельных точек сетчатки. На рис 34 представлена схема этого сложнейшего по своему строению аппарата. Наличие в нем элементов, обеспечивающих как центростремительную, так и центробежную организацию импульсов, позволяет видеть рефлекторный, фугально-петальный принцип строения этой периферической части зрительного анализатора. Таким образом, сетчатка глаза представляет собой аппарат, который не только получает извне определенные раздражения, но и регулируется прямыми влияниями центральных импульсов, которые изменяют возбудимость отдельных воспринимающих элементов и позволяют как дробить, так и объединять структуры возбуждения.
Рис. 34. Нейронная организация сетчатки (по Кригу). Обозначение слоев сетчатки: I и II — слой палочек и колбочек; ///—IV—V — слой биполярных клеток; VI — слой ганглионарных клеток. Обозначения видов клеток; am — амакринные клетки; с — внешние горизонтальные клетки; / — внутренние горизонтальные клетки; d — h — биполярные клетки; m—s — ганглионарные клетки; t—u — рецептивные части колбочек и палочек Возбуждения, возникающие в сетчатке глаза, по зрительным волокнам, частично перекрещивающимся в хиазме, передаются в центральные подкорковые и корковые нервные аппараты: в наружное коленчатое тело и ядра зрительного бугра и далее в зрительные отделы затылочной области коры головного мозга. До последнего времени считалось, что волокна зрительного нерва, зрительного тракта и той части зрительного пути, которая проходит внутри белого вещества больших полушарий и имеет название «зрительного сияния», являются лишь центростремительными, афферентными. Однако рядом исследований последнего времени (Поляк, 1957; Е. Г. Школьник-Яррос, 1958 и др.) было установлено, что все эти части зрительного пути имеют и значительное число центробежных, эфферентных волокон и что, таким образом, центральный аппарат зрения носит в полном смысле этого слова рефлекторный характер.
Как показывает схема, разработанная Е. Г. Школьник-Яррос (рис. 35), большинство идущих от сетчатки волокон, прерываясь в ядрах наружного коленчатого тела, направляется к той части затылочной коры, которая располагается в полюсе затылочной области и затем идет по ее медиальной поверхности. Эта часть (поле 17-е Бродмана) с полным основанием может быть оценена как первичное поле коркового ядра зрительного анализатора. Некоторое количество эфферентных волокон, прерываясь в наружном коленчатом теле и поступая затем во «вторичные» (вентральное и заднее) ядра зрительного бугра, доходит до тех отделов коры затылочной области (поле 18-е и 19-е Бродмана), которые могут быть обозначены как «широкая зрительная сфера» (Петцль, 1928) или как вторичные поля коркового ядра зрительного анализатора (см. выше I, 2) Из всех перечисленных полей зрительной коры берут начало как эфферентные волокна, направляющиеся к подкорковым зрительным ядрам, и от них к сетчатке, так и ассоциативные волокна, идущие от 17 поля к 18-му и 19-му и обратно. Все вместе они составляют тот рефлекторный элемент зрительной коры, который осуществляет сложные акты зрительного восприятия и патология которого нас интересует.
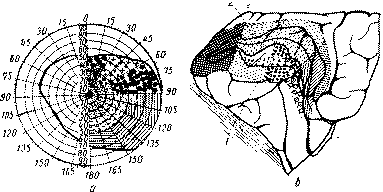
Рис. 36 Проекция сетчатки на кору (по Г. Холмсу). На рисунке изображено соответствие участков зрительной зоны коры (поля 17-го Бродмана) и участков зрительного поля: а — зрительное поле; б — зрительная зона коры (1 — нижняя губа шпорной борозды, 2 — верхняя губа шпорной борозды) Столь же существенным, как и фугально-петальное строение проводящего аппарата зрительной рецепции, является и топическая организация самой зрительной коры. Еще со времени Брауэра (1917—1932, сводное изложение 1936) хорошо известно, что волокна, идущие от сетчатки, располагаются в строгом порядке, так что отдельные части наружного коленчатого тела и проекционных отделов зрительной коры соответствуют определенным частям сетчатки (рис. 36). В силу этого раздражение соответствующих точек зрительной коры вызывает фотопсии в определенных частях зрительного поля, а разрушение этих участков зрительной коры приводит к строго определенному выпадению соответствующих (контралатеральных) участков зрительного поля, известных под названием гемианопсий и скотом. Тщательное исследование этих явлений было в последнее время проведено Тейбером (1960). К центральному или проекционному участку «коркового ядра» зрительного анализатора примыкают «вторичные зрительные поля», или поля «широкой зрительной сферы» (поля 18-е и 19-е). По своему цитоархитектоническому строению они отличаются от поля 17-го только тем, что четвертый афферентный слой клеток здесь суживается, уступая место более развитому, преимущественно ассоциативному, комплексу второго и третьего слоя Подобно вторичной слуховой коре, в 18-е и 19-е поля поступают волокна из тех ядер зрительного бугра, которые не имеют прямой связи с периферией и несут импульсы, уже подвергшиеся предварительной переработке в нижележащих подкорковых образованиях (Поляк, 1957 и др.). Таким образом, и эти зоны (аналогично 22-му полю височной коры) относятся к тем разделам, которые приобрели название «вторичных» корковых зон. Нейронографические исследования, проведенные Бонином, Гэрол и Мак-Кэлло-ком (1942) и др , показали, что этому архитектоническому строению вторичных полей затылочной области соответствуют определенные физиологические свойства. Как показывает рис. 37, при раздражении проекционных отделов зрительной коры (поля 17-го) возбуждение распространяется только в пределах данного поля, в то время как эффект раздражения 18-го поля носит гораздо более широкий характер и может быть прослежен как на соседних (и даже далеко отстоящих) полях, так и на полях противоположного полушария. Если верить данным указанных авторов, местное раздражение 19-го поля вызывает столь же широкий, но обратный по знаку тормозящий эффект.
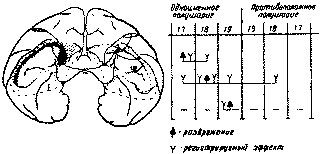
Рис. 37. Распространение возбуждения по коре при раздражении первичных и вторичных отделов затылочной области (по Мак-Кэллоку) Сходные данные были получены и при раздражении на операционном столе коры затылочной области человека. Исследования, начатые Хоффом и Петцлем (1930) и широко продолженные Пенфилдом (1941, 1954 и др.), показали, что раздражение проекционных отделов затылочной области приводит лишь к возникновению элементарных зрительных ощущений (цветные круги, туман, языки пламени и т. п.), в то время как раздражение вторичных отделов зрительной коры вызывает более сложные, содержательные зрительные галлюцинации с оживлением зрительных образов, сложившихся у больного в его предшествующем опыте. На рис. 38 мы приводим сводную схему тех точек коры, при раздражении которых возникали зрительные галлюцинации. Различный по своей сложности характер работы отдельных участков зрительной коры получает в этих опытах свое фактическое подтверждение.
Рис. 38. Схема точек затылочной области, раздражение которых вызывает зрительные галлюцинации (по Петцлю, Хоффу и др.). Цифрами обозначены те участки мозговой коры, при раздражении которых возникали соответствующие зрительные галлюцинации. Раздражения поля 17-го: 1 — «Светящиеся шары». 2 — «Окрашенный свет». 3 — «Белый свет». 4 — «Голубой диск». 5 — «Пламя». 6—«Голубой туман». Раздражения полей 18—19-го: 7 —«Лица, звуки, идущие со стороны вниз». 8 — «Человек, идущий со стороны». 9 — «Лица», 10 — «Звери». 11 — «Лица и бабочки» Роль коры затылочной области в анализе и синтезе зрительных раздражений была подробно исследована И. П. Павловым и его сотрудниками на собаках, Лешли (1930— 1942) на крысах, Клювером (Г927, 1937—1941) «а обезьянах. Этими исследованиями было показано, что если на низших этапах эволюции элементарные формы зрительного анализа и синтеза еще могут быть обеспечены подкорковыми образованиями, то уже у грызунов удаление коры затылочной области, оставляя сохранным различение общей массы света, устраняет всякую возможность различения форм. Еще более грубые нарушения зрительного анализа и синтеза возникают при экстирпации этих отделав коры у приматов. Однако наиболее существенный факт, показанный И. П. Павловым и его школой, заключается в том, что экстирпация коры затылочной области у собаки приводит не столько к снижению остроты зрения, сколько к нарушению более сложных форм анализа и синтеза зрительных сигналов. Поэтому, как хорошо известно из исследований этой школы, у собаки с экстирпированной затылочной областью могут вырабатываться условные рефлексы только на простые зрительные сигналы, но не образуются более сложные дифферанцировки, основанные на выделении более тонких зрительных признаков (форма, цвет и т. д.). Реакции на такие зрительные сигналы теряют свой избирательный характер. Именно это нарушение селективности составляет ту особенность, в которой, согласно учению И. П. Павлова, можно выразить функцию зрительной коры головного мозга. Близкие данные были получены в последнее время рядом американских исследователей (Чоу, 1952; Мишкин, 1954; Мишкин и Приб-рам, 1954; Орбах и Фанц, 1958 и др.). Эти авторы, работавшие на обезьянах, показали, что экстирпация тех отделов коры, которые являются гомологичными «вторичными» полями коркового ядра зрительного анализатора, приводит у них к заметному нарушению выработки сложных зрительных дифференцировок, хотя более элементарные зрительные функции остаются относительно сохранными. Факты, полученные на животных, представляют несомненный интерес для понимания общих принципов работы зрительной коры. Однако наиболее богатые данные открываются при наблюдении за теми функциональными изменениями, которые возникают при поражениях затылочной области мозговой коры у человека. |