Высшие корковые функции человека и их нарушения при локальных поражениях мозга
Александр Романович Лурия
Этапы пройденного пути
Научная автобиография

|
Высшие корковые функции человека и их нарушения при локальных поражениях мозгаАлександр Романович ЛурияЭтапы пройденного пути
|
|
|
|
|
|||
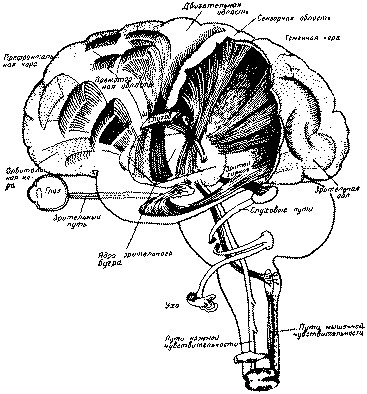
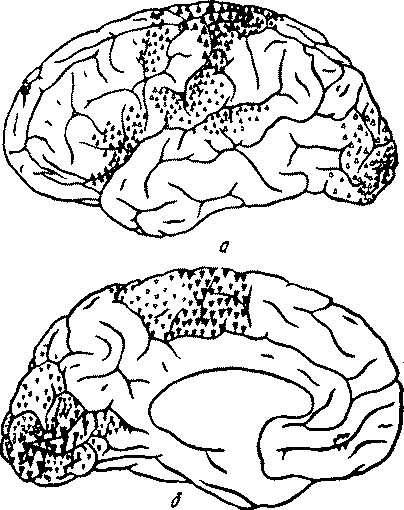
Высшие корковые функции человека и их нарушения при локальных поражениях мозгаб) Особенности структурной организации задних отделов корыФизиологическими экспериментами и клиническими наблюдениями, проведенными еще во второй половине XIX в., в задних отделах коры мозга были установлены области (рис. 7), в которые проецируются ре-цепторные поверхности как органов чувств, обращенных во внешний мир (зрения, слуха, кожной чувствительности), так и заложенные непосредственно в органах движения (кинестетический, или, употребляя выражение И. П. Павлова, двигательный анализатор). Дальнейшее накопление экспериментально-физиологического и клинического материала, особенно многочисленных случаев черепно-мозговых ранений, позволило более точно установить состав корковых полей ядерных зон анализаторов. Отправляясь от основного павловского определения ядерной зоны как такого отдела коры, который имеет дело с наиболее тонкими дифференцировками и наиболее сложными синтезами специальных раздражений, мы должны, следовательно, включать в состав ядерной зоны того или иного анализатора именно те поля, повреждение которых сопровождается, как правило, разными формами нарушения специальной функции данного анализатора. Таким образом, мы приходим к заключению, что затылочная область коры (поля 17, 18 и 19) является ядерной зоной зрительного анализатора, верхняя височная подобласть (поля 41, 42 и 22) —ядерной зоной слухового анализатора, а постцентральная область (поля 3, 1,2) — ядерной зоной кожно-кинестетического анализатора.
Рис. 7. Корковые зоны систем анализаторов (по Пейпецу, 1959) Многолетний опыт клинических наблюдений показывает, что повреждения различных полей, входящих в состав коркового ядра анализатора, отнюдь не равноценным образом сказывается на клинической картине. В этом отношении среди остальных полей выделяется одно, занимающее центральное положение в ядерной зоне. Таким центральным, или первичным, полем является в зрительной зоне поле 17, в слуховой — поле 41, в кожно-кинестетической— поле 3. Именно при повреждении этих центральных полей наступают резко выраженные симптомы выпадения способности непосредственно воспринимать и осуществлять» наиболее тонкие дифференцировки соответствующих раздражителей. Это обстоятельство (на котором мы еще будем останавливаться неоднократно) получило в последние годы дальнейшее подтверждение в экспериментах Пенфилда и Джаспера (1959), проведенных у операционного стола. При электрическом раздражении этих полей у больного возникают отдельные изолированные ощущения света и цвета или ощущения звука, или ощущения раздражения кожи и т. п.
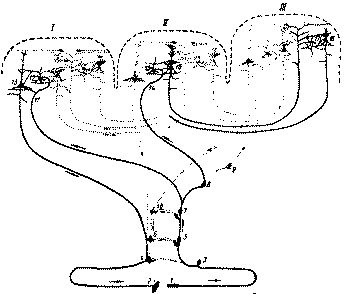
Рис. 8. Системы связей первичных, вторичных и третичных полей коры (по Г. И. Полякову): /—первичные (центральные) поля; // — вторичные (периферические) поля; III — третичные поля (зоны перекрытия анализаторов). Жирными линиями выделены: / — система проекционных (корково-подкорковых) связей коры; // — система проекционно-ассоциационных связей коры; III — система ассо-циационных связей коры. 1 — рецептор; 2 — эффектор; 3 — нейрон чувствительного узла; 4— двигательный нейрон; 5—6 — переключательные нейроны спинного мозга и ствола; 7—10 — переключательные нейроны подкорковых образований; 11, 14 — афферентное волокно из подкорки; 13 — пирамида V слоя; 16 — пирамида подслоя III3; 18 — пирамиды подслоев III2 и III1; 12, 15, 17 — звездчатые клетки коры Центральные поля ядерных зон резко выделяются своеобразием своей цитоархитектонической картины (пылевидная, или кониокорти-кальная, кора). Именно в этих полях особенно многочисленны мелкие зерновидные клетки слоя IV, переключающие импульсы, приходящие по мощно развитым афферентным проекционным волокнам из подкорковых . отделов анализаторов, на пирамидные нейроны слоев III и V корыЛ / В первичном зрительном поле 17 наряду с этими элементами, сгруппированными здесь в три подслоя слоя IV, обращают на себя внимание также очень крупные звездчатые клетки слоя IV и пирамидные клетки слоя V (так называемые клетки Кахала и клетки Майнерта), которые дают начало нисходящим проекционным волокнам, направляющимся к центрам движения глаз в среднем мозге. Все перечисленные особенности объединяют эти образования в первичный проекционный нейронный комплекс коры (рис. 8, I). Из этих особенностей нейронного строения первичных полей видно, что в этих полях относительно наибольшего развития достигает комплекс нейронов, (приспособленных для установления двусторонних корково-подкорковых связей, наиболее прямым и 'коротким иутем (т. е. с наименьшим числом переключений в подкорке) соединяющих кору с воспринимающими поверхностями соответствующих органов чувств (см. рис. 9).
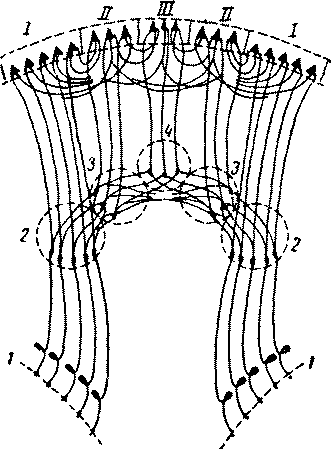
Рис 9 Схема переключений в системах анализаторов (по Г. И Полякову): / — периферические воспринимающие поверхности анализаторов; 2, 3, 4 — переключения в подкорковых отделах анализаторов. /, //, /// — как на рис. 8 Благодаря замыканию многочисленных как поступательных, так и обратных связей с нижележащими переключательными инстанциями анализаторов первичные поля коры в состоянии весьма подробно выделять отдельные раздражители, устанавливая рецепторы через посредство эф-фекторных приспособлений органов чувств на оптимальное восприятие соответствующих раздражителей. Это осуществляется с помощью замыкающихся в коре «собственных», или «местных», рефлексов данного анализатору. Важнейшей общей чертой структурной и функциональной организации первичных полей является наличие в них четкой соматотопической проекции, при которой отдельные точки периферии (кожной поверхности, скелетной мускулатуры тела, сетчатки глаза, улитки внутреннего уха) проецируются в строго отграниченные соответствующие точки первичных полей коры.По этой причине эти поля и получили название «проекционных» полей коры. Важно при этом отметить, что соматотопическая проекция воспринимающей поверхности на центральное корковое поле строится не по зеркально-геометрическому, а по функциональному принципу. Другими словами, отдельные участки тела представлены в первичных полях коры не пропорционально их величине, а пропорционально их физиологическому значению. Соответственно в центральной зрительной коре непропорционально большое место занимает область проекции центральной части сетчатки (желтого пятна), осуществляющей наиболее острое видение; в кожнокинестетической зоне относительно наибольшей протяженностью отличаются участки, куда проецируются кожные и мышечные рецепции пальцев и кисти, характеризующиеся наиболее выраженной способностью дискриминации отдельных мельчайших раздражителей (рис. 10). Ha основании всего вышесказанного ясно, что первичные поля ядерных зон представляют собой у человека высокоспециализированные участки коры, позволяющие весьма дробно выделять отдельные мельчайшие раздражители, что является совершенно необходимым условием синтетического восприятия сложных комплексов раздражителей, данных многообразием внешнего мира.
Рис 10 Схема соматотопической проекции в коре головного мозга (по Пенфилду) А — корковая проекция чувствительности, Б — корковая проекция двигательнойсистемы Относительные размеры органов отражают ту площадь коры головногомозга, с которой могут быть вызваны соответствующие ощущения и движения Наличие строго разграниченных отдельных пунктов проекций в пределах первичных полей следует понимать не как статическую, а как подвижную, динамическую конструкцию. Это особенно отчетливо выступает в первичном двигательном поле коры (прецентральное гиганто-пирамидное поле 4). Как мы уже говорили, еще в исследованиях О. Ферстера (1936), Грэм Брауна и Шеррингтона (1912) и других было показано, что электрическое раздражение, приложенное к одной и той же точке двигательной коры, может давать различные эффекты в зависимости от различной интенсивности прикладываемого тока и предшествующего состояния данного коркового участка. Например, при многократном раздражении центров мышц сгибателей сгибание может смениться разгибанием и т. д. Периферические, или вторичные, поля, расположенные в периферических отделах ядерных зон, характерным образом отличаются от центральных, или первичных, полей по своим физиологическим проявлениям и по особенностям архитектоники и нейронной организации. Эффекты, наступающие как при поражении, так и при электрическом раздражении этих полей, касаются преимущественно более сложных форм психических процессов. В случаях повреждения периферических полей ядерных зон при относительной сохранности элементарных ощущений расстраивается главным образом способность адекватно отражать целые комплексы и взаимные соотношения составных элементов воспринимаемых объектов. Раздражения периферических полей зрительной и слуховой коры электрическим током или факторами, вызывающими эпилептические припадки, сопровождаются развернутыми в определенной пространственно-временной последовательности зрительными и слуховыми галлюцинациями. К этим фактам мы еще вернемся ниже (см. II, 3, б). /Цитоархитектонически во вторичных полях ядерных зон коры выступают на первый план те элементы их нейронной структуры, которые приспособлены для переключения приходящих из подкорки афферентных импульсов через посредство зерновидных клеток слоя IV на крупные пирамиды нижнего уровня слоя III (рис. 11), дающие начало наиболее мощной системе ассоциаци-онных связей коры (вторичный проекционно-ассоциацион-ный нейронный комплекс, см. рис 8,11). Из этой характернейшей особенности конструкции вытекает, что вторичные поля ядерных зон играют весьма существенную роль в осуществлении взаимной связи отдельных раздражителей, выделяемых через посредство первичных полей, а также в функциональном объединении ядер ных зон разных анализаторов в процессе интеграции сложных комплексов рецепций, идущих по фазным анализаторам. Эти поля имеют, таким образом, преимущественное отношение к осуществлению более сложных форм координированных психических процессов, связанных с детальным анализом соотношений конкретных предметных раздражителей и с ориентировкой в конкретном окружающем пространстве и времени. Указанная здесь черта нейронной организации периферических полей ядерных зон находится в согласии с данными нейрофизиологических исследований. Как показали проведенные методом нейронографии эксперименты (Мак Кэллок, 1943), возбуждение, возникшее в результате раздражения первичных полей коры, имеет лишь сравнительно ограниченное распространение непосредственно в пределах соответствующих ядерных зон. Напротив, возбуждение, возникающее во вторичных полях, имеет тенденцию распространяться на значительно более широкие территории коры за пределами данной ядерной зоны (см. II, 3, б и II, 4, б).
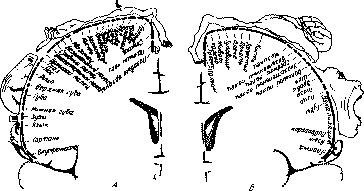
Рис 11 Распределение крупных клеток V и III слоев по
поверхности полушарий (видоизменено по Бейли и Бонину) Черные треугольники —
крупные клетки слоя V, светлые треугольники— крупные клетки подслоя III3 а —
наружная поверхность, б — внутренняя поверхность
В соответствии с большей сложностью устанавливаемых ассоциа-ционных связей проекционные связи вторичных полей коры характеризуются по сравнению с первичными большей сложностью всей системы переключений в подкорковых инстанциях анализаторов (см. рис. 9). Афферентные импульсы, направляющиеся в кору от рецепторных поверхностей органов чувств, достигают этих полей через большее число дополнительных переключений в так называемых ассоциационных ядрах зрительного бугра, в отличие от афферентных импульсов, направляющихся в первичные поля коры более коротким путем через так называемые реле-ядра зрительного бугра. Таким образом, рецепции, достигающие периферических полей и используемые для широких ассоциаций как внутри соответствующих зон, так и между разными зонами коры уже в подкорковых уровнях анализаторов, проходят определенную степень предварительной интеграции, которая распространяется не только на систему данного анализатора, но и на разные анализаторы, кооперирующиеся между собой в процессе координированной совместной деятельности. Морфологическим выражением дальнейшего усложнения функциональных отношений между анализаторами является тот факт, что в процессе прогрессивного роста ядерных зон анализаторов по поверхности полушария возникает взаимное перекрытие этих зон с образованием особых «зон перекрытия» корковых концов анализаторов (третичные поля коры, рис. 12). Эти корковые формации, представленные в задней части полушария полями верхней и нижней теменной областей, средней височной и височно-теменно-затылочной подобластей, имеют отношение к наиболее сложным формам интеграции совместной деятельности зрительного, слухового и кожно-кинестетического анализаторов. Поскольку третичные поля коры находятся уже за пределами собственных ядерных зон, их повреждение или раздражение не сопровождается выраженными явлениями выпадения или эффектами со стороны специфических функций анализаторов. Как мы увидим ниже, при поражении этих территорий расстраиваются главным образом наиболее обобщенные проявления корковой деятельности, опирающ иеся на совместную работу нескольких анализаторов, позволяющие осуществлять наиболее сложные формы ориентировки во внешнем мире и анализ и синтез сложных систем отношений между раздражителями, воздействующими на воспринимающую часть разных анализаторов. Многочисленные группы нейронов, из которых образованы третичные поля коры, по сравнению с комплексами нейронов ядерных зон, представляются почти полностью освобожденными от специфических анализаторных функций, переключаясь целиком на адекватное отражение наиболее сложных форм пространственных и временных отношений между реальными комплексами раздражителей и на оперирование этими отношениями в процессе активного воздействия организма на внешний мир. В этом плане можно рассматривать третичные поля коры как собрание «рассеянных» элементов анализаторов в павловском понимании этого термина, т. е. таких элементов, которые уже не способны сами по себе производить сколько-нибудь сложные анализы и синтезы специальных раздражителей. Богатый клинический опыт учит, однако, что эти «рассеянные» элементы анализаторов в максимальной степени концентрируют в себе потенциальные возможности функциональных взаимодействий между анализаторами, протекающих притом на высшем уровне интегративной деятельност и коры. Обширная у человека территория, занимаемая зонами перекрытия корковых концов анализаторов, распадается на ряд полей. Особенности микроструктуры этих полей, так же как и их функциональное значение, определяются топографическими соотношениями этих полей с полями ядерных зон, между которыми они располагаются. Оправданным представляется выделение трех частных зон перекрытия, представленных соответственно верхней теменной областью, нижней теменной областью и височно-теменно-затылочной подобластью. Обе теменные области топографически и функционально наиболее тесно связаны с зонами кожно-кинестетического и зрительного анализаторов, между которыми эти области располагаются.
Рис 12. Схема корково-подкорковых соотношений первичных, вторичных и третичных зон (по Г. И. Полякову) Жирными линиями показаны системы анализаторов с их переключениями в подкорковых отделах: / — зрительный анализатор; 2 — слуховой анализатор; 3 — кожно-кинестетический анализатор Τ — височная область; О — затылочная область; Pip — поле 39; Pia — поле 40; Pstc — постцентральная область; ТРО — височно-теменно-затылочная подобласть; Th.—зрительный бугор; Cgm—внутреннее коленчатое тело; Cgi — наружное коленчатое тело Верхняя теменная область граничит спереди с теми разделами постцентральной области, которые заняты представительством ног и туловища. Эта область соответственно имеет специальное значение для интеграции со зрительными рецепциями движений всего тела и в формировании схемы тела, нарушения которой были специально изучены в клинике (П. Шильдер, 1935 и др.). Нижняя теменная область граничит с теми разделами постцентральной области, которые заняты представительством рук и лица. Соответственно эта область имеет отношение к интеграции обобщенных и отвлеченных форм сигнализации, которые связаны с тонко и сложно дифференцированными предметными и речевыми действиями, совершаемыми под контролем арения и требующими совершенно разработанной системы ориентировки в окружающем пространстве. Наконец, височно-теменно-затылочная подобласть, составляющая зону перехода между слуховой и зрительной зонами коры, как показывают клинические наблюдения, функционально особенно тесно связана с наиболее сложными формами интеграции слуховых и зрительных рецепций, и специально, как мы увидим ниже (см. 11, 2 и 3), с определенной стороной семантики устной и письменной речи. Именно в пределах этой подобласти и составляющей ее продолжение кпереди средней височной извилины Пенфилд и Джаспер (1959) вызывали зрительные и слуховые галлюцинации при раздражении отдельных пунктов этой территории коры электрическим током. В архитектонической и нейронной картине зон перекрытия на первый план выступает третичный ассоциационный комплекс коры (см. рис. 8, III), представленный пирамидными и звездчатыми клетками наиболее высоких по уровню расположения и наиболее поздно дифференцирующихся в эволюции млекопитающих слоев и подслоев коры — среднего и верхнего подслоев слоя III и слоя II. Нейроны этой части серого вещества коры посылают, принимают и переключают наиболее тонко и сложно дифференцированные потоки импульсов, посредством которых осуществляется функциональное объединение нейронов ядерных зон в бесконечно разнообразные и подвижные рабочие констелляции в соответствии с характером осуществляемых высших психических функций. Наряду с весьма дробно дифференцированными многосторонними ассоциационными связями третичные поля коры характеризуются по сравнению с первичными и вторичными полями и наиболее сложными цепями проекционных переключений в подкорковых отделах анализаторов. В соответствии с наиболее высоким уровнем обобщения и отвлечения от конкретных реальных раздражителей, корковые фармации, о которых здесь идет речь, связаны с периферическими концами анализаторов наиболее опосредованным образом: через наибольшее число последовательных переключений в подкорковых образованиях (см. рис. 9). Корковые «зоны перекрытия» устанавливают двусторонние связи с наиболее поздно дифференцирующимся в эволюции комплексом ядер зрительного бугра (подушка зрительного бугра), которые связаны с собственно переключательными ядрами зрительного бугра («реле-ядрами») через сложную цепь внутренних переключений в самом зрительном бугре. Таким образом, импульсы, подаваемые снизу к этим территориям коры, проходят наиболее сложную переработку в подкорке и подводятся к коре, как интегрированные сигналы, образовавшиеся в результате взаимодействия чувственных показаний различных анализаторов. Суммируя все вышесказанное, следует особенно подчеркнуть, что в нормально действующем мозге описанные выше три группы корковых полей вместе с их системами переключений и связей между собой и с подкорковыми инстанциями анализаторов работают как одно сложно-дифференцированное целое. |