Большая Советская Энциклопедия
(БСЭ)
|
|
|
|
источник статьи: Большая Советская Энциклопедия (БСЭ) |
|
Трансформация в генетике, внесение в клетку генетической информации при помощи изолированной дезоксирибонуклеиновой кислоты (ДНК). Т. приводит к появлению у трансформированной клетки (трансформанта) и её потомства новых признаков, характерных для объекта — источника ДНК. Явление Т. было открыто в 1928 английским учёным Ф. Гриффитом, наблюдавшим наследуемое восстановление синтеза капсульного полисахарида у пневмококков при заражении мышей смесью убитых нагреванием капсулированных бактерий и клеток, лишённых капсулы. Организм мыши в этих экспериментах играл роль своеобразного детектора, так как приобретение капсульного полисахарида сообщало клеткам, лишённым капсулы, способность вызывать смертельный для животного инфекционный процесс (см. схему). В последующих экспериментах было установлено, что Т. имеет место и в том случае, когда вместо убитых клеток к лишённым капсулы пневмококкам добавляли экстракт из разрушенных капсулированных бактерий. В 1944 О. Эйвери с сотрудниками (США) установил, что фактором, обеспечивающим Т., являются молекулы ДНК. Эта работа — первое исследование, доказавшее роль ДНК как носителя наследственной информации. Помимо пневмококков, Т. обнаружена и изучена на некоторых других бактериях. Использование в экспериментах легко учитываемых генетических признаков (например, устойчивость к действию клеточных ядов, потребность в определённых факторах роста), а также применение ДНК с радиоизотопной меткой позволили дать Т. количественную оценку. Т. у бактерий рассматривают как сложный процесс, включающий следующие стадии: фиксация молекул ДНК клеткой-реципиентом; проникновение ДНК внутрь клетки; включение фрагментов трансформирующей ДНК в хромосому клетки-хозяина; формирование «чистых» трансформированных вариантов. Фиксация ДНК происходит на особых участках клеточной поверхности (рецепторах), число которых ограничено. Связанная с рецепторами ДНК сохраняет чувствительность к действию добавленного в среду фермента дезоксирибонуклеазы, вызывающего её распад. Однако, спустя очень короткий срок (в пределах 1 мин) после фиксации, часть ДНК проникает в клетку. Бактериальные клетки одного и того же штамма резко различаются по проницаемости для ДНК. Клетки данной бактериальной популяции, способные включать чужеродную ДНК, называются компетентными. Число компетентных клеток в популяции незначительно и зависит от генетических особенностей бактерий и фазы роста бактериальной культуры. Развитие компетенции связывают с синтезом особого белка, обеспечивающего проникновение ДНК в клетку. Средние размеры фрагментов ДНК, проникающих в клетку, составляют 5×106 дальтон. Поскольку в компетентную клетку может одновременно проникнуть ряд таких фрагментов, суммарная величина поглощённой ДНК может быть примерно равна размерам хромосомы клетки-хозяина. После проникновения в клетку двунитевой ДНК одна нить распадается до моно- и олигонуклеотидов, вторая — встраивается в хромосому клетки-хозяина путём её разрывов и воссоединений. Последующая репликация такой гибридной структуры приводит к выщеплению «чистых» клонов трансформантов, в потомстве которых закреплен признак, кодируемый включившейся ДНК. Применение Т. позволило провести генетический анализ бактерий, у которых не описано иных форм генетического обмена (конъюгации, трансдукции). Кроме того, Т. — удобный метод для выяснения влияний на биологическую активность ДНК физических или химических изменений её структуры. Разработка метода Т. у кишечной палочки позволила использовать для Т. не только фрагменты бактериальной хромосомы, но и ДНК бактериальных плазмид и бактериофагов. Этот метод широко используется для внесения в клетку гибридной ДНК в исследованиях по так называемой генной инженерии. Имеются сообщения о воспроизведении Т. на клетках высших организмов. Однако в этом случае процесс Т. изучен недостаточно.
Лит.: Хэйс У., Генетика бактерий и бактериофагов, пер. с англ., М., 1965; Прозоров А. А., Генетическая трансформация у микроорганизмов, М., 1966; Браун В., Генетика бактерий, пер. с англ., М., 1968; Бреслер С. Е., Молекулярная биология, Л., 1973; Стент Г., Молекулярная генетика, пер. с англ., М., 1974, гл. 7. А. Л. Табачник. 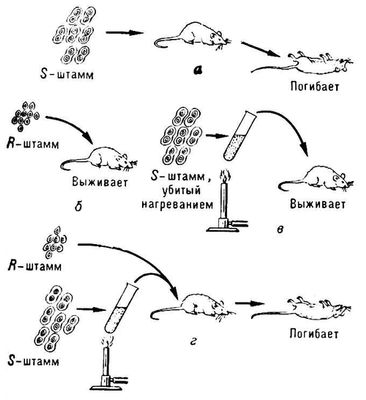 Схема эксперимента Гриффита (по Стенту): а — мышь, которой введена культура патогенного капсулированного штамма S пневмококов, погибает; б — мышь, которой введена культура непатогенного бескапсульного R—мутанта нормального S—штамма, не погибает; в — мышь, которой введена культура S—штамма, убитого предварительно нагреванием, не погибает; г — мышь, которой введена смесь живой культуры R—мутанта и убитой нагреванием культуры нормального S—штамма, погибает; в этом случае присутствие убитых нагреванием S—бактерий вызвало трансформацию живых R—бактерий, в результате чего у них восстановилась способность к образованию капсулы и патогенность. |