Большая Советская Энциклопедия
(БСЭ)
|
|
|
|
источник статьи: Большая Советская Энциклопедия (БСЭ) |
|
Мутации (от лат. mutatio — изменение, перемена), внезапно возникающие естественные (спонтанные) или вызываемые искусственно (индуцированные) стойкие изменения наследственных структур живой материи, ответственных за хранение и передачу генетической информации. Способность давать М. — мутировать — универсальное свойство всех форм жизни от вирусов и микроорганизмов до высших растений, животных и человека; оно лежит в основе наследственной изменчивости в живой природе. М., возникающие в половых клетках или спорах (генеративные М.), передаются по наследству; М., возникающие в клетках, не участвующих в половом размножении (соматические мутации), приводят к генетическому мозаицизму: часть организма состоит из мутантных клеток, другая — из немутантных. В этих случаях М. могут наследоваться только при вегетативном размножении с участием мутантных соматических частей организма (почек, черенков, клубней и т. п.). Внезапное возникновение наследственных изменений отмечалось многими учёными 18 и 19 вв., было хорошо известно Ч. Дарвину, но углублённое изучение М. началось лишь с зарождением на пороге 20 в. экспериментальной генетики. Термин «М.» ввёл в генетику в 1901 Х. Де Фриз. Типы мутаций. По характеру изменения генетического аппарата М. делят на геномные, хромосомные и генные, или точковые. Геномные М. заключаются в изменении числа хромосом в клетках организма. К ним относятся: полиплоидия — увеличение числа наборов хромосом, когда вместо обычных для диплоидных организмов 2 наборов хромосом их может быть 3, 4 и т. д.; гаплоидия — вместо 2 наборов хромосом имеется лишь один; анеуплоидия — одна или несколько пар гомологических хромосом отсутствуют (нуллисомия) или представлены не парой, а лишь одной хромосомой (моносомия) либо, напротив, 3 или более гомологичными партнёрами (трисомия, тетрасомия и т. д.). К хромосомным М., или хромосомным перестройкам, относятся: инверсии — участок хромосомы перевёрнут на 180°, так что содержащиеся в нём гены расположены в обратном порядке по сравнению с нормальным; транслокации — обмен участками двух или более негомологичных хромосом; делеции — выпадение значительного участка хромосомы; нехватки (малые делеции) — выпадение небольшого участка хромосомы; дупликации — удвоение участка хромосомы; фрагментации — разрыв хромосомы на 2 части или более. Генные М. представляют собой стойкие изменения химического строения отдельных генов и, как правило, не отражаются на наблюдаемой в микроскоп морфологии хромосом. Известны также М. генов, локализованных не только в хромосомах, но и в некоторых самовоспроизводящихся органеллах цитоплазмы (например, в митохондриях, пластидах; см. Наследственность цитоплазматическая). Изменения признаков организма, вызываемые мутациями. В результате М. могут изменяться самые различные биохимические, физиологические и морфологические признаки организма. Изменения эти у организмов, претерпевших М., — мутантов — могут быть резко выраженными или слабыми, представляющими лишь незначительные отклонения от среднего для данного вида значения признака (см. илл.). Полиплоидные мутанты обычно характеризуются увеличением размеров клеток и всего организма. Если у полиплоида число наборов хромосом чётное (сбалансированные полиплоиды), то плодовитость обычно сохраняется или понижена не сильно; полиплоиды же, у которых число наборов хромосом нечётное (несбалансированные полиплоиды), бесплодны или обладают низкой плодовитостью (при созревании половых клеток хромосомы распределяются в них беспорядочно, что приводит к образованию анеуплоидных гамет, большей частью неспособных к оплодотворению или дающих нежизнеспособные зиготы). Гаплоидные мутанты имеют мелкие клетки, размеры организма уменьшены по сравнению с диплоидной нормой, наблюдается полное или почти полное бесплодие, т. к. лишь немногие гаметы содержат полный набор хромосом. Анеуплоиды характеризуются весьма значительными изменениями различных признаков организма, нередко столь сильными, что вызывают его гибель или бесплодие. Обычно менее резкие изменения наблюдаются в случае делеций, нехваток и дупликаций, причём степень изменения признаков в общем пропорциональна длине выпавшего или удвоенного участка хромосомы (крупные делеции могут вызывать гибель организма). Инверсии и транслокации сами по себе не вызывают изменений признаков организма (если не сопровождаются эффектом положения гена, т. е. изменением его фенотипического проявления вследствие соседства с иными, чем прежде, генами), но приводят к существенным генетическим последствиям, т. к. у гетерозигот по инверсиям затруднён обмен участками между нормальной и несущей инверсию хромосомой (см. Кроссинговер), а гетерозиготы по транслокациям дают частично анеуплоидные, часто нежизнеспособные, половые клетки. Это же происходит в случае фрагментации в результате утери фрагмента хромосомы, оставшегося без центромеры. Генные М., составляющие основную долю всех М., вызывают чрезвычайно разнообразные изменения признаков организма, причём изменение одного гена обычно приводит к изменению нескольких признаков (см. Плейотропия). Генные М. могут быть доминантными, полудоминантными и рецессивными (см. Доминантность, Рецессивность). В результате М. ген может переходить в разные состояния (множественные аллели одного и того же гена), по-разному влияющие на контролируемые данным геном признаки организма. Мутантные гены могут отличаться от соответствующих нормальных тем, что специфический для данного гена продукт (чаще всего фермент) не образуется вовсе; образуется в меньшем или превышающем норму количестве; образуется продукт, инактивирующий или тормозящий продукт немутантного гена; вместо нормального образуется иной, не взаимодействующий с ним продукт, отсутствующий у немутантных особей. Претерпевший М. ген обычно столь же стабилен, как немутантный, из которого он произошёл; вследствие новой М. он может вернуться к исходному состоянию (обратные М.). Генные М., как правило, вредны для организма, они нарушают жизненные процессы, протекающие в организме, снижают его жизнеспособность и плодовитость; нередко мутантный ген обусловливает гибель развивающегося организма (летальные М.). Реже возникают генные М., сравнительно мало влияющие на жизнеспособность и плодовитость организма, ещё реже — улучшающие те или иные его свойства. Эта последняя категория генных М., несмотря на свою относительную редкость, имеет огромное значение, т. к. даёт основной материал как для естественного отбора, так и для искусственного отбора, являясь необходимым условием эволюции и селекции. Причины мутаций и их искусственное вызывание. Полиплоидия чаще возникает, когда хромосомы в начале клеточного деления — митоза — разделились, но деления клетки почему-либо не произошло. Искусственно полиплоидию удаётся вызвать, воздействуя на вступившую в митоз клетку веществами, нарушающими цитотомию. Реже полиплоидия бывает следствием слияния 2 соматических клеток или участия в оплодотворении яйцеклетки 2 спермиев. Гаплоидия — большей частью следствие развития зародыша без оплодотворения (см. Партеногенез). Искусственно её вызывают, опыляя растения убитой пыльцой или пыльцой др. вида (отдалённого). Основная причина анеуплоидии — случайное нерасхождение пары гомологичных хромосом при мейозе, в результате чего обе хромосомы этой пары попадают в одну половую клетку или в неё не попадает ни одна из них. Реже возникают анеуплоиды из немногих оказавшихся жизнеспособными половых клеток, образуемых несбалансированными полиплоидами. Причины хромосомных перестроек и наиболее важной категории М. — генных — долгое время оставались неизвестными. Это давало повод для ошибочных автогенетических концепций (см. Автогенез), согласно которым спонтанные генные М. возникают в природе якобы без участия воздействий окружающей среды. Лишь после разработки методов количественного учёта генных М. выяснилась возможность вызывать их различными физическими и химическими факторами — мутагенами. Первые данные о влиянии излучений радия на наследственную изменчивость у низших грибов были получены в СССР (Г. А. Надсон и Г. С. Филиппов, 1925). Убедительные доказательства возможности искусственно вызывать М. были приведены в 1927 Г. Мёллером, обнаружившим в опытах на дрозофиле сильное мутагенное действие рентгеновских лучей. В дальнейшем работами по генетическому действию излучений на различные организмы была установлена универсальная способность всех ионизирующих излучений вызывать не только генные М., но и хромосомные перестройки. Мутагенное действие некоторых химических веществ было впервые обнаружено в СССР М. Н. Мейселем (1928), В. В. Сахаровым (1933) и М. Е. Лобашёвым (1934); первый сильный химический мутаген (чужеродная ДНК) был открыт в 1939 С. М. Гершензоном с сотрудниками; в 1946 сильное мутагенное действие формалина и этиленимина было установлено советским генетиком И. А. Рапопортом, иприта — английскими генетиками Ш. Ауэрбах и Д. Робсоном. Позже были открыты сотни других химических мутагенов. Сильные физические и химические мутагены увеличивают частоту возникновения генных М. и хромосомных перестроек во много десятков раз, а наиболее мощные химические мутагены (так называемые супермутагены, многие из которых открыты и изучены советским генетиком И. А. Рапопортом с сотрудниками) — даже в сотни раз по сравнению с частотой возникающих естественно спонтанных М. В опытах на культурах клеток и на лабораторных животных обнаружено мутагенное действие многих вирусов. Мутагеном у вирусов, по-видимому, служит их нуклеиновая кислота. Т. о., вирусы — не только возбудители многих болезней животных и человека, растений и микроорганизмов, но и один из источников их наследственной изменчивости. Все мутагены вызывают генные М., прямо или косвенно изменяя молекулярную структуру нуклеиновых кислот, в которой закодирована генетическая информация. Экспериментальные исследования спонтанных и индуцированных М. (наиболее изучены М. у кукурузы, дрозофилы, а также ряда микроорганизмов) вскрыли ряд важных особенностей мутирования генов. Частота возникновения спонтанных М. неодинакова для разных генов и различных организмов, составляя для отдельного гена от 1:105 до 1:107 в поколение; немногие, так называемые мутабильные, гены характеризуются значительно более высокой частотой мутирования. Частота прямых и обратных М. одного и того же гена нередко различна. Мутагены повышают частоту М. примерно одинаково для всех генов, так что соотношение более часто и сравнительно редко мутирующих генов («спектр» М.) остаётся приблизительно одинаковым как при спонтанном, так и при индуцированном мутационным процессе (в случае химических мутагенов могут наблюдаться небольшие различия в спектрах вызываемых ими М.). Лишь у микроорганизмов некоторые химические мутагены сильнее повышают частоту мутирования определённых генов, чем остальных («горячие точки» хромосом). Сходное явление обнаружено при мутагенном действии нуклеиновых кислот и вирусов на многоклеточные организмы. Соотношение общего числа генных М. и хромосомных перестроек различно при действии физических и химических мутагенов — для вторых характерна большая доля генных М., чем для первых; те или иные различия имеются и в действии разных химических мутагенов. Далеко не все изменения, вызываемые мутагенами в ДНК клетки, реализуются в М. Во многих случаях поврежденный участок ДНК удаляется в процессе рекомбинации или «вырезается» имеющимися в клетке так называемыми репарирующими ферментами, восстанавливающими структуру ДНК, и при дальнейшей репликации ДНК замещается соответствующим нормальным участком (см. Репарация). Частота любых М. зависит от многих внешних и внутренних факторов — температуры, парциального давления кислорода, возраста организма, фазы развития и физиологического состояния клетки и др. Большое значение имеют особенности генотипа: даже в пределах одного вида генетически разнящиеся линии могут обладать различной мутабильностью. У ряда организмов описаны так называемые гены-мутаторы, резко повышающие частоту М. Благодаря зависимости мутабильности от генетических факторов, её удаётся повышать или понижать искусственным отбором. Неодинаковая мутабильность разных видов — следствие аналогичного действия естественного отбора в ходе их эволюции. Значение мутаций для эволюции, селекции и медицины. Основы понимания роли М. в эволюции были заложены в 20-х гг. 20 в. работами советского генетика С. С. Четверикова, английских учёных Дж. Холдейна и Р. Фишера и американского учёного С. Райта, положивших начало развитию эволюционной генетики. Было показано, что все наследственные изменения, служащие материалом для эволюции, обязаны М. (комбинативная изменчивость, возникающая путём образования новых сочетаний генов при скрещивании, в конечном счёте, тоже есть следствие М., обусловливающих генетические различия скрещивающихся особей). В отличие от модификаций, М. не являются однозначной реакцией на вызывающее их воздействие: один и тот же мутагенный фактор приводит к возникновению разнообразных М., затрагивающих те или иные признаки организма и изменяющих их в разных направлениях. Поэтому сами по себе М. не имеют адаптивного характера. Однако постоянно возникающие у любого вида живых существ М., многие из которых к тому же длительно сохраняются в популяции в скрытом виде (рецессивные М.), служат резервом наследственной изменчивости, который позволяет естественному отбору перестраивать наследственные признаки вида, приспосабливая его к меняющимся условиям среды (изменению климата или биоценоза, переселению в новый ареал и т. п.). Т. о., адаптивность эволюционных изменений — следствие сохранения естественным отбором носителей тех М. и их сочетаний, которые оказываются полезными в данной обстановке. При этом М., бывшие в одних условиях вредными или нейтральными, могут оказаться полезными в изменившихся условиях. Наибольшее значение для эволюции имеют генные М. Несмотря на относительную редкость М. каждого гена, общая частота спонтанных генных М. весьма значительна, т. к. генотип многоклеточных организмов состоит из десятков тысяч генов. В результате ту или иную генную М. несёт большая доля образуемых организмом гамет или спор (у высших растений и животных эта доля достигает 5—30%), что создаёт предпосылки для эффективного действия естественного отбора. Хромосомные перестройки, затрудняющие рекомбинацию, — инверсии и транслокации — способствуют репродуктивной изоляции отдельных групп организмов и их последующей дивергенции (см. Видообразование); дупликации ведут к увеличению числа генов в генотипе и возрастанию их разнообразия вследствие происходящей затем дифференциации генов в дуплицированных участках хромосом. Полиплоидия играет большую роль в эволюции растений; при этом, помимо репродуктивной изоляции, она в ряде случаев восстанавливает плодовитость бесплодных межвидовых гибридов. С разработкой способов искусственного мутагенеза открылась возможность значительного ускорения селекции — селекционерам стал доступен гораздо больший исходный материал, чем при использовании редких спонтанных мутаций. В 1930 советские учёные А. А. Сапегин и Л. Н. Делоне впервые применили ионизирующую радиацию в селекции пшеницы. В дальнейшем методами радиационной селекции были выведены новые высокоурожайные сорта пшеницы, ячменя, риса, люпина и др. с.-х. растений, ценные штаммы микроорганизмов, используемых в промышленности. В селекции с хорошими результатами применяются и химические мутагены. Геномные М., хромосомные перестройки и генные М. — причина многих наследственных заболеваний и врождённых уродств у человека. Поэтому ограждение человека от действия мутагенов — важнейшая задача. Огромное значение в этом отношении имело осуществлённое по инициативе СССР запрещение испытаний ядерного оружия в атмосфере, загрязняющих окружающую среду радиоактивными веществами. Очень важно тщательное соблюдение мер защиты человека от радиации в атомной индустрии, при использовании радиоактивных изотопов, рентгеновских лучей и т. п. Необходимо изучение возможного мутагенного действия различных новых лекарственных средств, пестицидов, химических препаратов, применяемых в промышленности, и запрещение производства тех из них, которые окажутся мутагенными. Профилактика вирусных инфекций имеет значение и для защиты потомства от мутагенного действия вирусов. См. также Генетика, Генетика микроорганизмов, Изменчивость, Молекулярная генетика, Радиобиология.
Лит.: Супермутагены. Сб. ст., М., 1966; Лобашев М. Е., Генетика, 2 изд., Л., 1967, гл. 11, 14; Гершкович И., Генетика, пер. с англ., М., 1968, гл. 11—14, 30, 31; Сойфер В. Н., Молекулярные механизмы мутагенеза, М., 1969; Дубинин Н. П., Общая генетика, М., 1970, гл. 17, 20; Ратнер В. А., Принципы организации и механизмы молекулярно-генетических процессов, Новосиб., 1972, гл. 3; Serra J. A., Modern genetics, v. 3, L.-N. Y., 1968, ch. 20—22; Auerbach C., Kilbey B. J., Mutation in Eukaryotes, «Annual Review of Genetics», 1971, v. 5, p. 163; Banks G. R., Mutagenesis: a review of some molecular aspects, «Science Progress», 1971, v. 59, № 236. С. М. Гершензон.
 Мутантные формы ячменя: поздняя полегающая (слева) и ранняя неполегающая (справа).  Соматические мутации, вызванные у растений ионизирующей радиацией (рентгеновские или гамма-лучи): появление белой окраски в красных цветках табака (1) и двух сортов львиного зева (2 и 3); на рис. 3 (слева) — нормальный цветок, справа — мутировавший после облучения. 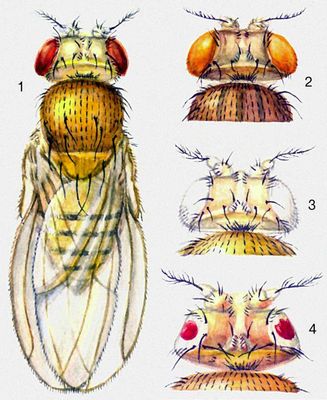 Мутации окраски и формы глаз у плодовой мушки — дрозофилы: 1 — дикий тип — тускло-красные глаза; мутантные формы: 2 — розовые глаза, 3 — белые глаза, 4 — уменьшенные, «плосковидные».  Мутации окраски шерсти у домовой мыши: 1 — дикий тип — серая окраска; мутантные формы: 2 — белая, 3 — желтая, 4 — чёрная, 5 — коричневая, 6 — мелкокрапчатая.  Мутации окраски у канареек: 2 — дикий тип — зелёная; мутантные формы: 1 — жёлтая, 3 — пятнистая. |